Аннотированных учебных элементов. Aллель - вариант (состояние) гена, локализованного в определенном локусе (месте) хромосомы
Сцепленное наследование. Г. Мендель опубликовал результаты своих исследований в 1865 г., однако тогда его открытия остались незамеченными. Только в 1900 г. К-Корренс (Германия), Г. де Фриз (Голландия) и Э. Чер мак (Австрия) независимо друг от друга обнаружили у разных видов растений те же закономерности наследования признаков, что и Г. Мендель. Английский генетик У. Бэтсон подтвердил законы Менделя на животных. Переоткрытие законов Менделя вызвало глубокий интерес к изучению закономерностей наследования признаков и способствовало быстрому развитию генетики.
В 1902 г. немецкий цитолог и эмбриолог Т. Б о в е р и представил доказательства участия хромосом в процессах передачи наследственной информации. Он показал, например, что нормальное развитие морского ежа возможно лишь при наличии всех хромосом. Подобную связь заметил в 1903 г. и американский цитологУ. С эттон. Так получили обоснование предположения Менделя
о наследственных факторах, о наличии одинарного набора этих факторов в гаметах и двойного - в зиготах. В 1909 г. датский биолог В. Иогансен ввел понятие s.ген:/.
В 1910 г. американский генетик Т. Морган экспериментально доказал, что гены расположены в хромосомах. Многочисленные исследования Моргана и его учеников привели к целому ряду важнейших открытий, которые легли в основу хромосомной теории наследственности. Одно из ее положений можно сформулировать следующим образом: гены расположены в хромосомах в линейном порядке и занимают определенные участки - локусы, причем аллельные гены находятся в одинаковых локусах гомологичных хромосом.
Закон независимого наследования (третий закон Менделя) справедлив в том случае, если неаллельные гены находятся в разных парах хромосом. Однако количество генов у живых организмов значительно больше числа хромосом. Например, у человека около 25 тыс. генов, а количество хромосом -
23 пары (2п = 46); у плодовой мушки дрозофилы приблизительно 14 тыс. генов и всего 4 пары хромосом (2п = 8). Следовательно, каждая хромосома содержит множество генов. Будут ли гены, локализованные в одной хромосоме, наследоваться независимо? Очевидно, что нет.
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Совместное наследование генов Т. Морган предложил называть сцепленным наследованием (в отличие от независимого). Каждая пара гомологичных хромосом содержит гены, контролирующие одни и те же признаки, поэтому количество групп сцепления равно числу пар хромосом. Например, у человека 23 группы сцепления, а у дрозофилы - 4.
Вам известно, что при независимом наследовании дигетерозиготная особь, например =^=, образует четыре типа гамет в равном соотношении, т. е. по 25 %: Л В, АЬ, а В и ab. Это обусловлено тем, что неаллельные гены находятся в разных парах хромосом. Если же они расположены в гомологичных хромосомах, следовало бы ожидать, что дигетерозигота будет производить лишь два типа га- ab
мет: 50 % АВ и 50 % ab (обратите внимание на то, что сцепленные гены записываются в одну хромосому).
Однако Т. Морган обнаружил, что в большинстве случаев дигетерозиготные особи образуют не два, а четыре типа гамет. Помимо ожидаемых АВ и ab формируются также гаметы с новыми комбинациями генов: АЬ и аВ, только в меньшем процентном соотношении. Рассмотрим один из экспериментов Т. Моргана, в котором изучалось наследование сцепленных генов у дрозофилы.
Если почистить фрукты или овощи и не сразу выбросить очистки либо оставить фрукты на столе на несколько дней, то можно заметить, как вокруг остатков пищи начнут роиться маленькие мушки размером около 2-3,5 мм. Это дрозофилы - плодовые мушки, род насекомых отряда Двукрылые (рис. 95). Обычно дрозофилы имеют красные глаза и желтокоричневую окраску брюшка. Жизненный цикл дрозофил короток: развитие от яйца до половозрелой особи при 25 °С занимает 10 дней. Небольшие размеры, высокая плодовитость, простота культивирования и ряд других особенностей на долгое время сделали дрозофилу главным объектом генетики. Не один нобелевский лауреат, кроме своего интеллекта, обязан ей своими научными достижениями.
Путем скрещивания чистой линии дрозофил, имеющих серое тело и нормальные (длинные) крылья, с чистой линией, особи которой имели черное тело и зачаточные крылья, были получены гибриды первого поколения (рис. 96). Все они в соответствии с законом единообразия были серыми с нормально развитыми крыльями. Следовательно, у дрозофил серое тело (А) полностью доминирует над черным (а), а нормальные крылья (В) - над зачаточными (b ). Все гибриды первого поколения - дигетерозиготы.
Затем было проведено анализирующее скрещивание (рис. 97). Дигетерозиготную самку из гибридного поколения скрестили с рецессивным дигомозиготным самцом (черное тело и зачаточные крылья). В потомстве было получено по 41,5 % особей с серым телом, нормальными крыльями и черным телом, зачаточными крыльями, а также по 8,5 % мух с серым телом, зачаточными крыльями и черным телом, нормальными крыльями.
Если бы гены, определяющие цвет тела и развитие крыльев, находились в разных парах хромосом, соотношение фенотипических классов было бы равным - по 25 %. Но этого не наблюдалось, значит, гены находятся в гомологичных хромосомах и наследуются сцепленно.
Несмотря на сцепление генов, АВ самка производила не два, а четыре типа гамет. Однако гамет с исходными сочетаниями сцепленных генов формировалось намного больше (АВ и ab вместе составили 83 %), чем с новыми их сочетаниями (сумма АЬ и дВ равна 17 %).
Было выяснено, что причиной появления хромосом с новыми комбинациями родительских генов является кроссинговер. Вы помните, что этот процесс происходит в профазе I мейоза и представляет собой обмен соответствующими участками между гомологичными хромосомами. Таким образом, кроссинговер препятствует полному (абсолютному) сцеплению генов. Гаметы, которые образуются в результате кроссинговера, и особи, которые развиваются при участии таких гамет, называются кроссоверными или рекомбинантными. В рассмотренном эксперименте гаметы АЬ и аВ являлись кроссоверными, а гаметы АВ и ab - некроссоверными (см. рис. 97).
Кроссинговер между конкретными сцепленными генами происходит с определенной вероятностью (частотой). Для расчета частоты кроссинговера (rf, от англ. recombination frequency - частота рекомбинации) можно пользоваться следующей формулой:
Таким образом, между генами А и В, контролирующими цвет тела и длину крыльев дрозофилы, кроссинговер происходит с частотой: rf AB = 17 %.
Дальнейшие исследования, проведенные Т. Морганом и его сотрудниками, показали, что частота кроссинговера пропорциональна расстоянию между генами, расположенными в одной хромосоме. Чем больше расстояние между сцепленными генами, тем чаще между ними происходит кроссинговер. И наоборот, чем ближе друг к другу расположены гены, тем меньше частота кроссинговера между ними. Чем объясняется эта закономерность?
В профазе I мейоза при конъюгации гомологичных хромосом образование перекрестов между хроматидами осуществляется произвольно, на любых соответствующих участках. Рассмотрим рисунок 98.
Гены А и В (или а. и Ь) находятся сравнительно близко друг к другу. Вероятность того, что перекрест произойдет именно на участке, разделяющем эти гены, невелика. Гены А и D (или а. и d) располагаются на значительном расстоянии друг от друга. Поэтому вероятность того, что хроматиды перекрестятся на каком-либо участке между ними, намного выше. Значит, чем больше расстояние между генами, тем чаще они разделяются при кроссинговере.
Таким образом, частота кроссинговера позволяет судить о расстоянии между генами. В честь Т. Моргана единица измерения расстояния между генами получила название моргай и да или, что то же самое, санти моргай и да (сМ).
Морганида (сантиморганида, сМ) - это генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Биологическое значение кроссинговера чрезвычайно велико. В результате этого процесса возникают новые комбинации родительских генов, что повышает генетическое разнообразие потомства и расширяет возможности адаптации организмов к различным условиям окружающей среды.
Генетические карты. Т. Морган и сотрудники его лаборатории показали, что знание частоты кроссинговера между сцепленными генами позволяет строить генетические карты хромосом. Генетическая карта представляет собой схему взаимного расположения генов, находящихся в одной группе сцепления, с учетом расстояний между ними (рис. 99).
Генетические карты хромосом уже составлены для человека, многих видов животных, растений, грибов и микроорганизмов. Наличие генетической карты свидетельствует о высокой степени изученности того или иного вида организма и представляет большой научный интерес. Такой организм является прекрасным объектом для проведения дальнейших экспериментальных работ, имеющих не только научное, но и практическое значение. В частности, знание генетических карт позволяет планировать работы по получению организмов с определенными сочетаниями признаков, что широко применяется в селекционной практике. Генетические карты хромосом человека используются в медицине для диагностики и лечения ряда наследственных заболеваний.
Основные положения хромосомной теории наследственности.
1. Гены в хромосомах расположены линейно, в определенной последовательности. Аллельные гены находятся в одинаковых локусах гомологичных хромосом.
2. Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Количество групп сцепления равно числу пар хромосом.
3. Сцепление генов может нарушаться в результате кроссинговера, происходящего при конъюгации гомологичных хромосом в профазе I мейоза.
4. Частота кроссинговера пропорциональна расстоянию между генами: чем больше расстояние, тем выше частота кроссинговера, и наоборот.
Б. За единицу расстояния между сцепленными генами принята 1 морганида - расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Формы взаимодействия неаллельных генов (межаллельное взаимодействие)
Эти гены могут располагаться в разных локусах гомологичных хромосом или в негомологичных хромосомах, обычно отвечают за развитие разных признаков.
Комплементарность (лат. комплементум – дополнение) - присутствие в одном генотипе двух доминантных (рецессивных) генов, которые дополняют действие друг друга, и признак формируется лишь при одновременном действии обоих генов.
Пример: развитие слуха у человека. Для нормального слуха в генотипе человека должны присутствовать доминантные гены из разных аллельных пар –D и E. Ген D отвечает за нормальное развитие улитки, а ген E – за развитие слухового нерва. У рецессивных гомозигот (dd) будет недоразвита улитка, а при генотипе (ее) недоразвит слуховой нерв. Люди с генотипами D..ee, ddE.. и ddee будут глухими.
Эпистаз – такой вид взаимодействия, при котором доминантный (рецессивный) ген из одной аллельной пары подавляет действие доминантного (рецессивного) гена из другой аллельной пары. Соответственно эпистаз может быть как доминантным таки рецессивным. Это явление противоположно комплементарности. Подавляющий ген называется супрессором, ингибитором, эпистатичным. Подавляемый ген – гипостатичным. У человека описан «бомбейский феномен» в наследовании групп крови по АВО системе. У женщины получившей от матери аллель J B фенотипически определялась I(О) группа крови. При детальном исследовании было установлено, что действие гена J B было подавлено редким рецессивным геном, который в гомозиготном состоянии оказал эпистатическое действие.
Полимерия – доминантные гены из разных аллельных пар влияют на степеньпроявления одного и тогоже признака. Полимерные гены принято обозначать одной буквой латинского алфавита с цифровыми индексами. Так у человека количество пигмента меланина в коже (и, следовательно, цвет кожи) определяется четырьмя неаллельными генами: Р 1 - Р 4 . Соответственно темно-коричневый цвет кожи будут иметь люди с генотипом Р 1 Р 1 Р 2 Р 2 Р 3 Р 3 Р 4 Р 4 . Самому светлому цвету кожи соответствует генотип р 1 р 1 - р 4 р 4 . Промежуточные варианты будут определять различную интенсивность пигментации: Например, человек с большим количеством доминантных генов в генотипе будет иметь более темную кожу. Признаки, детерминируемые полимерными генами, называются полигенными, для них свойственен большой диапазон изменчивости, т.е. широкая норма реакции. Таким образом, наследуются многие количественные и некоторые качественные признаки – рост, масса тела, величина артериального давления.
Основные закономерности наследования признаков по Менделю реализуются благодаря существованию закона (гипотезы) чистоты гамет , выдвинутого Г. Менделем в 1865г.
Суть последнего состоит в том, что пара аллельных генов, определяющая тот или иной признак: а) никогда не смешивается; б) в процессе гаметогенеза расходится в разные гаметы, то есть в каждую из них попадает один ген из аллельной пары. Цитологически это обеспечивается мейозом: аллельные гены лежат в гомологичных хромосомах, которые в анафазе мейоза расходятся к разным полюсам и попадают в разные гаметы.
II. Дигибридное скрещивание
Ранее мы изучали закономерности наследования 1 признака (моногибридное скрещивание)
В общей и медицинской генетике часто возникает необходимость в изучении одновременного наследования двух или более признаков (ди- и полигибридное скрещивание). Если каждый их этих признаков контролируются парой аллельных генов, то можно предположить существование двух форм наследования: независимого и сцепленного. Принципиальные отличия будут определяться расположением генов в хромосомах. При сцепленном наследовании обе пары аллельных генов располагаются в одной паре гомологичных хромосом (т.е. в одной группе сцепления). При независимом наследовании пары аллельных генов располагаются в разных парах гомологичных хромосом.
Закономерности и механизмы независимого наследования были выявлены и сформулированы Г.Менделем в 3-м законе «Закон независимого комбинирования признаков»: при скрещивании гомозиготных организмов, отличающихся по двум (или более) парам альтернативных признаков, в первом поколении наблюдается единообразие по гено- и фенотипу, а при скрещивании гибридов первого поколения – во втором наблюдается расщепление по фенотипу 9:3:3:1, и при этом возникают организмы с комбинациями признаков, не свойственных родительским формам».
Для этой цели Мендель использовал гомозиготные растения гороха, отличающиеся по двум парам альтернативных признаков: семена желтые, гладкие и зеленые, морщинистые. В первом скрещивании он получил АаВb растения с желтыми, гладкими семенами, т.е закон единообразия гибридов первого поколения проявляется не только при моногибридном, но и полигибридном скрещивании, если родительские формы гомозиготны.
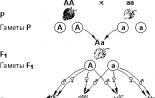
P: ААВВ х ааbb
G: АВ, АВ аb, ab
F 1 : АаВb
P (F 1 ): АаВb х АаВb
|
ААВВ |
АаВВ | |||
|
АаВВ |
ааВВ | |||
F 2 : 9: 3: 3: 1
9 частей растений с горошинами желтыми, гладкими, три части с желтыми морщинистыми, 3 с зелеными гладкими, и 1часть с зелеными, морщинистыми, (3+1) n - расщепление по фенотипу, где n- число анализируемых признаков.
Возникают организмы с новыми комбинациями признаков, не свойственных родительским формам.
Условия выполнения закона:
Признаки наследуются моногенно (наследование по каждой паре идет независимо)
Форма взаимодействия аллельных генов – полное доминирование
Пары аллельных генов располагаются в разных парах гомологичных хромосом
У человека независимо наследуются цвет глаз и цвет волос.
Причины разнообразия гибридов:
Независимое расхождение пар хромосом в анафазу I мейоза (приводит к образова-
нию гамет с различными комбинациями неаллельных генов)
Случайное слияние гамет при оплодотворении (возникают различные комбинации
генов в генотипах потомков, которые определяют комбинацию признаков)
Новые комбинации генов в генотипах потомков приводят к возникновению у них новых комбинаций признаков – главный вывод 3-го закона.
В 1908г. Сэттон и Пеннет обнаружили отклонения от свободного комбинирования признаков согласно IIIзакону Менделя. В 1911-12г. Т.Морган с сотр. Описалиявление сцепления генов – совместную передачу группы генов из поколения в поколение.
У дрозофилы гены окраски тела (b+ - серое тело,b– черное тело) и длины крыльев (vg+ - нормальные крылья,vg– короткие крылья), находятся в одной хромосоме, это сцепленные гены находящиеся в одной группе сцепления. Если скрестить двух гомозиготных особей с альтернативными признаками, то в первом поколении, все гибриды будут иметь одинаковый фенотип с проявлениями доминантных признаков (серое тело, нормальные крылья).
Это не противоречит закону единообразия гибридов Iпоколения Г.Менделя. Однако при дальнейшем скрещивании гибридов первого поколения между собой вместо ожидаемого расщепления по фенотипу 9:3:3:1, при сцепленном наследовании происходило расщепление в отношении 3:1, появились особи только с признаками родителей, а особей с перекомбинацией признаков не было.
Это связано с тем, что в мейозе гаметогенеза к полюсам клетки расходятся целые хромосомы. Одна хромосома из данной гомологичной пары и все гены, которые находятся в ней, отходят к одному полюсу и в дальнейшем попадают в одну гамету. Другая хромосома из данной пары отходит к противоположному полюсу и попадает в другую гамету. Совместное наследование генов находящихся в одной хромосоме, называется сцепленным наследованием.
Примером полного сцепления генов у человека может служить наследование резус фактора. Наличие резус-фактора обусловлено тремя сцепленными меду собой генами, поэтому наследование его происходит по типу моногибридного скрещивания.
Однако гены, находящиеся в одной хромосоме, иногда могут наследоваться раздельно, в этом случае говорят о неполном сцеплении генов
Продолжая свои работы по дигибридному скрещиванию, Морган провел два опыта по анализирующему скрещиванию и выявил, что сцепление генов может быть полным и неполным.
Причина неполного сцепления генов – кроссинговер. В мейозе при конъюгации гомологичные хромосомы могут перекрещиваться и обмениваться гомологичными участками. В этом случае гены одной хромосомы переходят в другую, гомологичную ей.
В период роста гаметогенеза происходит редупликация ДНК, генетическая характеристика овоциов и сперматоцитов Iпорядка 2n4c, каждая хромосома состоит из двух хроматид, которые содержат идентичный набор ДНК. В профазу редукционного деленя мейоза происходит коньюгация гомологичных хромосом и может произойти обмен аналогичными участками гомологичных хромосом –кроссинговер. В анафазу редукционного деления к полюсам расходятся целые гомологичные хромосомы, после завершения деления образуются клеткиn2c– овоциты и сперматоцитыIIпорядка. В анафазу эквационного деления расходятся хроматиды –nc, но при этом они отличаются комбинацией неаллельных генов.Новые комбинации неаллельных генов – генетический эффект кроссинговера. → новые комбинации признаков у потомков → комбинативная изменчивость.
Чем ближе друг к другу расположены гены в хромосоме, тем сильнее между ними сцепление и тем реже происходит их расхождение при кроссинговере, и, наоборот, чем дальше друг от друга отстоят гены, тем слабее сцепление между ними и тем чаще возможно его нарушение.
полное сцепление Схема кроссинговера
Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах: единице расстояния между генами, находящимися в одной хромосоме, соответствует 1% кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид.
Теоретической основой Закономерностей сцепленного наследования являются положенияХромосомной теории наследственности , которая была сформулирована и экспериментально доказана Т. Морганом и его сотрудниками в1911г. Ее сущность заключается в следующем:
Основным материальным носителем наследственности являются хромосомы с локализованными в них генами;
Гены расположены в хромосомах в линейном порядке в определенных локусах, аллельные гены занимают одинаковые локусы гомологичных хромосом.
Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются преимущественно вместе (или сцеплено); число групп сцепления равно гаплоидному набору хромосом.
Во время гаметогенеза (профаза I мейоза) может происходить обмен аллельными
генами - кроссинговер, который нарушает сцепления генов.
Частота кроссинговера пропорциональна расстоянию между генами. 1морганида – единица расстояния, равная 1% кроссинговера.
Данная теория дала объяснение законам Менделя, вскрыла цитологические основы наследования признаков.
Явление сцепления генов лежит в основе составления генетических карт хромосом – схемы относительного положения генов, находящихся в одной группе сцепления. Методы картирования хромосом направлены на то, чтобы узнать в какой хромосоме, и в каком ее локусе (месте) расположен ген, а также определить расстояние между соседними генами
Это отрезок прямой, на котором обозначен порядок расположения генов и указано расстояние между ними в морганидах, строится по результатам анализирующего скрещивания. Чем чаще признаки наследуются вместе, тем ближе гены, отвечающие за эти признаки, располагаются в хромосоме. Другими словами, о расположении генов в хромосоме можно судить по особенностям проявления признаков в фенотипе.
При анализе сцепления генов у животных и растений используется гибридологический метод, у человека – генеалогический метод, цитогенетический, а также метод гибридизации соматических клеток.
Цитологическая карта хромосомы представляет собой фотографию или точный рисунок хромосомы, на котором отмечается последовательность расположения генов. Ее строят на основе сопоставления результатов анализирующего скрещивания и хромосомных перестроек.
Лекц и я № 3
Хромосомная теория наследственности.
Основные положення хромосомной теории наследственности. Хромосомный анализ.
Формирование хромосомной теории. В 1902-1903 гг. американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бовери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911-1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Хромосомная теория наследственности - теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, то есть преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Хромосомная теория наследственности возникла в начале 20 в. на основе клеточной теории и использовалась для изучения наследственных свойств организмов гибридологического анализа.
Основные положения хромосомной теории наследственности.
1. Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
2. Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
3. Гены расположены в хромосоме в линейной последовательности.
4. Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
5. Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
6. Каждый биологический вид характеризуется определенным набором хромосом - кариотипом .
Сцепленное наследование
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы - более 1 тыс., а у человека - около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несколько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.
Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием . Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например, у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин - 24).
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов- Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер .
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость , которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе .
Сцепление и кроссинговер. Из принципов генетического анализа, изложенных в предыдущих главах, с очевидностью вытекает, что независимое комбинирование признаков может осуществляться лишь при условии, что гены, определяющие эти признаки, находятся в негомологичных хромосомах. Следовательно, у каждого организма число пар признаков, по которым наблюдается независимое наследование, ограничено числом пар хромосом. С другой стороны, очевидно, что число признаков и свойств организма, контролируемых генами, чрезвычайно велико, а число пар хромосом у каждого вида относительно мало и постоянно.
Остается предположить, что в каждой хромосоме находится не один ген, а много. Если это так, то третий закон Менделя касается распределения хромосом, а не генов, т. е. его действие ограничено.
Явление сцепленного наследования . Из третьего закона Менделя следует, что при скрещивании форм, различающихся двумя парами генов (АВ и а b ), получается гибрид А a В b , образующий четыре сорта гамет АВ, А b , аВ и а b в равных количествах.
В соответствии с этим в анализирующем скрещивании осуществляется расщепление 1: 1: 1: 1, т.е. сочетания признаков, свойственные родительским формам (АВ и а b ), встречаются с такой же частотой, как и новые комбинации (А b и аВ),- по 25%. Однако по мере накопления фактов генетики все чаще стали сталкиваться с отклонениями от независимого наследования. В отдельных случаях новые комбинации признаков (А b и аВ) в F b совсем отсутствовали - наблюдалось полное сцепление между генами исходных форм. Но чаще в потомстве в той или иной степени преобладали родительские сочетания признаков, а новые комбинации встречались с меньшей частотой, чем ожидается при независимом наследовании, т.е. меньше 50%. Таким образом, в данном случае гены чаще наследовались в исходном сочетании (были сцеплены), но иногда это сцепление нарушалось, давая новые комбинации.
Совместное наследование генов, ограничивающее их свободное комбинирование, Морган предложил называть сцеплением генов или сцепленным наследованием.
Кроссинговер и его генетическое доказательство. При допущении размещения в одной хромосоме более одного гена встает вопрос, могут ли аллели одного гена в гомологичной паре хромосом меняться местами, перемещаясь из одной гомологичной хромосомы в другую. Если бы такой процесс не происходил, то гены комбинировались бы только путем случайного расхождения негомологичных хромосом в мейозе, а гены, находящиеся в одной паре гомологичных хромосом, наследовались бы всегда сцепленно - группой.
Исследования Т.Моргана и его школы показали, что в гомологичной паре хромосом регулярно происходит обмен генами. Процесс обмена идентичными участками гомологичных хромосом с содержащимися в них генами называют перекрестом хромосом или кроссинговером. Кроссинговер обеспечивает новые сочетания генов, находящихся в гомологичных хромосомах. Явление кроссинговера, так же как и сцепление, оказалось общим для всех животных, растений и микроорганизмов. Наличие обмена идентичными участками между гомологичными хромосомами обеспечивает обмен или рекомбинацию генов и тем самым значительно увеличивает роль комбинативной изменчивости в эволюции. О перекресте хромосом можно судить по частоте возникновения организмов с новым сочетанием признаков. Такие организмы называют рекомбинантами.
Гаметы с хромосомами, претерпевшими кроссинговер, называют кроссоверными, а с непретерпевшими - некроссоверными. Соответственно организмы, возникшие от сочетания кроссоверных гамет гибрида с гаметами анализатора, называют кроссоверами или рекомбинантами, а возникшие за счет некроссоворных гамет гибрида - некроссоверными или нерекомбинантными.
Закон сцепления Моргана. При анализе расщепления в случае кроссинговера обращает на себя внимание определенное количественное отношение кроссоверных и некроссоверных классов. Обе исходные родительские комбинации признаков, образовавшиеся из некроссоверных гамет, оказываются в потомстве анализирующего скрещивания в равном количественном отношении. В указанном опыте с дрозофилой тех и других особей было примерно по 41,5%. В сумме некроссоверные мухи составили 83% от общего числа потомков. Два кроссоверных класса по числу особей также одинаковы, и сумма их равна 17%.
Частота кроссинговера не зависит от аллельного состояния генов, участвующих в скрещивании. Если в качестве родителя использовать мух и , то в анализирующем скрещивании кроссоверные (b + vg и bvg + ) и некроссоверные (bvg и b + vg + ) особи появятся с той же частотой (соответственно 17 и 83%), что и в первом случае.
Результаты этих опытов показывают, что сцепление генов реально существует, и лишь в известном проценте случаев оно нарушается вследствие кроссинговера. Отсюда и был сделан вывод, что между гомологичными хромосомами может осуществляться взаимный обмен идентичными участками, в результате чего гены, находящиеся в этих участках парных хромосом, перемещаются из одной гомологичной хромосомы в другую. Отсутствие перекреста (полное сцепление) между генами представляет исключение и известно лишь у гетерогаметного пола немногих видов, например у дрозофилы и шелкопряда.
Изученное Морганом сцепленное наследование признаков получило название закона сцепления Моргана. Поскольку рекомбинация осуществляется между генами, а сам ген кроссинговером не разделяется, его стали считать единицей кроссинговера.
Величина кроссинговера . Величина кроссинговера измеряется отношением числа кроссоверных особей к общему числу особей в потомстве от анализирующего скрещивания. Рекомбинация происходит реципрокно, т.е. между родительскими хромосомами осуществляется взаимный обмен; это обязывает подсчитывать кроссоверные классы вместе как результат одного события. Величина кроссинговера выражается в процентах. Один процент кроссинговера составляет единицу расстояния между генами.
Линейное расположение генов в хромосоме. Т. Морган предположил, что гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме; чем реже кроссинговер, тем они ближе друг к другу.
Одним из классических опытов Моргана на дрозофиле, доказывающим линейное расположение генов, был следующий. Самки, гетерозиготные по трем сцепленным рецессивным генам, определяющим желтую окраску тела y , белый цвет глаз w и вильчатые крылья bi , были скрещены с самцами, гомозиготными по этим трем генам. В потомстве было получено 1,2% мух кроссоверных, возникших от перекреста между генами у и w ; 3,5% − от кроссинговера между генами w и bi и 4,7% - между у и bi .
Из этих данных с очевидностью вытекает, что процент перекреста является функцией расстояния между генами. Поскольку расстояние между крайними генами у и bi равно сумме двух расстояний между у и w , w и bi , следует предположить, что гены расположены в хромосоме последовательно, т.е. линейно.
Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов в хромосоме строго фиксировано, т. е. каждый ген занимает в хромосоме свое опрделенное место - локус.
Основным положениям хромосомной теории наследственности - парности аллелей, их редукции в мейозе и линейному расположению генов в хромосоме - соответствует однонитчатая модель хромосомы.
Одинарный и множественный перекресты. Приняв положения, что генов в хромосоме может быть много и расположены они в хромосоме в линейном порядке, а каждый ген занимает определённый локус в хромосоме, Морган допустил, что перекрест между гомологичными хромосомами может происходить одновременно в нескольких точках. Это предположение было им доказано тоже на дрозофиле, а затем полностью подтвердилось на ряде других животных, а также на растениях и микроорганизмах.
Кроссинговер, происходящий лишь в одном месте, называют одинарным, в двух точках одновременно – двойным, в трёх – тройным и т.д., т.е. он может быть множественным.
Чем дальше отстоят друг от друга в хромосоме гены, тем больше вероятность двойных перекрестов между ними. Процент рекомбинаций между двумя генами тем точнее отражает расстояние между ними, чем оно меньше, так как в случае малого расстояния уменьшается возможность двойных обменов.
Для учета двойного кроссинговера необходимо иметь дополнительный маркер, находящийся между двумя изучаемыми генами. Определение расстояния между генами осуществляют следующим образом: к сумме процентов одинарных кроссоверных классов прибавляют удвоенный процент двойных кроссинговеров. Удвоение процента двойных кроссинговеров необходимо в связи с тем, что каждый двойной кроссинговер возникает благодаря двум независимым одинарным разрывам в двух точках.
Интерференция. Установлено, что кроссинговер, происшедший в одном месте хромосомы, подавляет кроссинговер в близлежащих районах. Это явление носит название интерференции. При двойном перекресте интерференция проявляется особенно сильно в случае малых расстояний между генами. Разрывы хромосом оказываются зависимыми друг от друга. Степень этой зависимости определяется расстоянием между происходящими разрывами: по мере удаления от места разрыва возможность другого разрыва увеличивается.
Эффект интерференции измеряется отношением числа наблюдаемых двойных разрывов к числу возможных при допущении полной независимости каждого из разрывов.
Локализация гена. Если гены расположены в хромосоме линейно, а частота кроссинговера отражает расстояние между ними, то можно определить местоположение гена в хромосоме.
Прежде чем определить, положение гена, т. е. его локализацию, необходимо определить, в какой хромосоме находится данный ген. Гены, находящиеся в одной хромосоме и наследующиеся сцепленно, составляют группу сцепления. Очевидно, что количество групп сцепления у каждого вида должно соответствовать гаплоидному набору хромосом.
К настоящему времени группы сцепления определены у наиболее изученных в генетическом отношении объектов, причем во всех этих случаях обнаружено полное соответствие числа групп сцепления гаплоидному числу хромосом. Так, у кукурузы (Zea mays ) гаплоидный набор хромосом и число групп сцепления составляют 10, у гороха (Pisum sativum ) – 7, дрозофилы (Drosophila melanogaster) – 4, домовой мыши (Mus musculus ) – 20 и т. п.
Поскольку ген занимает определенное место в группе сцепления, это позволяет устанавливать порядок расположения генов в каждой хромосоме и строить генетические карты хромосом.
Генетические карты. Генетической картой хромосом называют схему относительного расположения генов, находящихся в данной группе сцепления. Они составлены пока лишь для некоторых наиболее изученных с генетической точки зрения объектов: дрозофилы, кукурузы, томатов, мыши, нейроспоры, кишечной палочки и др.
Генетические карты составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют.
Для того, чтобы составить карты, необходимо изучить закономерности наследования большого числа генов. У дрозофилы, например, изучено более 500 генов, локализованных в четырех группах сцепления, у кукурузы - более 400 генов, локализованных в десяти группах сцепления и т.д. При составлении генетических карт указывается группа сцепления, полное или сокращенное название генов, расстояние в процентах от одного из концов хромосомы, принятого за нулевую точку; иногда обозначается место центромеры.
У многоклеточных организмов рекомбинация генов бывает реципрокной. У микроорганизмов она может быть односторонней. Так, у ряда бактерий, например у кишечной палочки (Escherichia coli ), перенос генетической информации происходит во время конъюгации клеток. Единственная хромосома бактерии, имеющая форму замкнутого кольца, рвется во время конъюгации всегда в определенной точке и переходит из одной клетки в другую.
Длина переданного участка хромосомы зависит от длительности конъюгации. Последовательность генов в хромосоме оказывается постоянной. В силу этого расстояние между генами на такой кольцевой карте измеряется не в процентах кроссинговера, а в минутах, что отражает продолжительность конъюгации.
Цитологическое доказательство кроссинговера. После того как генетическими методами удалось установить явление кроссинговера, необходимо было получить прямое доказательство обмена участками гомологичных хромосом, сопровождающегося рекомбинацией генов. Наблюдаемые в профазе мейоза картины хиазм могут служить лишь косвенным доказательством этого явления, констатация происшедшего обмена прямым наблюдением невозможна, так как обменивающиеся участками гомологичные хромосомы обычно абсолютна одинаковы но величине и форме.
Чтобы сопоставить цитологические карты гигантских хромосом с генетическими, Бриджес предложил воспользоваться коэффициентом кроссинговера. Для этого он разделил общую длину всех хромосом слюнных желез (1180 мкм) на общую длину генетических карт (279 единиц). В среднем это отношение оказалось равным 4,2. Следовательно, каждой единице перекреста на генетической карте соответствует 4,2 мкм на цитологической карте (для хромосом слюнных желез). Зная расстояние между генами на генетической карте какой-либо хромосомы, можно сравнить относительную частоту перекреста в разных ее районах. Например, в Х- хромосоме дрозофилы гены у и ec находятся на расстоянии 5,5%, следовательно, расстояние между ними в гигантской хромосоме должно быть 4,2 мкм Х 5,5 = 23 мкм, но непосредственное измерение дает 30 мкм. Значит, в этом районе Х -хромосомы кроссинговер идет реже средней нормы.
В силу неравномерного осуществления обменов по длине хромосом гены при нанесении их на карту распределяются на ней с разной плотностью. Следовательно, распределение генов на генетических картах можно рассматривать как показатель возможности осуществления перекреста по длине хромосомы.
Механизм кроссинговера. Еще до открытия перекреста хромосом генетическими методами цитологи, изучая профазу мейоза, наблюдали явление взаимного обвивания хромосом, образования ими χ-образных фигур – хиазм (χ-греческая буква «хи»). В 1909 г. Ф.Янсенс высказал предположение, что хиазмы связаны с обменом участками хромосом. Впоследствии эти картины послужили дополнительным аргументом в пользу гипотезы генетического перекреста хромосом, выдвинутой Т.Морганом в 1911 г.
Механизм перекреста хромосом связан с поведением гомологичных хромосом в профазе I мейоза.
Кроссинговер происходит на стадии четырех хроматид и приурочен к образованию хиазм.
Если в одном биваленте произошел не один обмен, а два и более, то и этом случае образуется несколько хиазм. Поскольку в биваленте четыре хроматиды, то, очевидно, каждая из них имеет равную вероятность обменяться участками с любой другой. При этом в обмене могут участвовать две, три или четыре хроматиды.
Обмен внутри сестринских хроматид не может приводить к рекомбинациям, поскольку они генетически идентичны, и в силу этого такой обмен не имеет смысла в качестве биологического механизма комбинативной изменчивости.
Соматический (митотический) кроссинговер. Как уже говорилось, кроссинговер происходит в профазе I мейоза при образовании гамет. Однако существует соматический, или митотический, кроссинговер, который осуществляется при митотическом делении соматических клеток главным образом эмбриональных тканей.
Известно, что гомологичные хромосомы в профазе митоза обычно не конъюгируют и располагаются независимо друг от друга. Однако иногда удается наблюдать синапсис гомологичных хромосом и фигуры, похожие на хиазмы, но при этом редукции числа хромосом не наблюдается.
Гипотезы о механизме кроссинговера. По поводу механизма перекреста существует несколько гипотез, но ни одна из них не объясняет полностью фактов рекомбинации генов и наблюдаемых при этом цитологических картин.
Согласно гипотезе, предложенной Ф.Янсенсом и развитой К.Дарлингтоном, в процессе синапсиса гомологичных хромосом в биваленте создается динамическое напряжение, возникающее в связи со спирализацией хромосомных нитей, а также при взаимном обвивании гомологов в биваленте. В силу этого напряжения одна из четырех хроматид рвется. Разрыв, нарушая равновесие в биваленте, приводит к компенсирующему разрыву в строго идентичной точке какой-либо другой хроматиды этого же бивалента. Затем происходит реципрокное воссоединение разорванных концов, приводящее к кроссинговеру. Согласно этой гипотезе хиазмы непосредственно связаны с кроссинговером.
По гипотезе К.Сакса хиазмы не являются результатом кроссинговера: сначала образуются хиазмы, а затем происходит обмен. При расхождении хромосом к полюсам вследствие механического напряжения в местах хиазм происходят разрывы и обмен соответствующими участками. После обмена хиазма исчезает.
Смысл другой гипотезы, предложенной Д.Беллингом и модернизированной И.Ледербергом, заключается в том, что процесс репликации ДНК может реципрокно переключаться с одной нити на другую; воспроизведение, начавшись на одной матрице, с какой-то точки переключается на матричную нить ДНК.
Факторы, влияющие на перекрест хромосом. На кроссинговер влияет множество факторов как генетической природы, так и внешней среды. Поэтому в реальном эксперименте о частоте кроссинговера можно говорить, имея в виду все те условия, в которых она была определена. Кроссинговер практически отсутствует между гетероморфными Х - и Y -хромосомами. Если бы он происходил, то хромосомный механизм определения пола постоянно разрушался бы. Блокирование кроссинговера между этими хромосомами связано не только с различием в их величине (оно наблюдается не всегда), но и обусловлено Y -специфичными нуклеотидными последовательностями. Обязательное условие синапса хромосом (или их участков) - гомология нуклеотидных последовательностей.
Для абсолютного большинства высших эукариот характерна примерно одинаковая частота кроссинговера как у гомогаметного, так и гетерогаметного полов. Однако есть виды, у которых кроссинговер отсутствует у особей гетерогаметного пола, в то время как у особей гомогаметного пола он протекает нормально. Такая ситуация наблюдается у гетерогаметных самцов дрозофилы и самок шелкопряда. Существенно, что частота митотического кроссинговера у этих видов у самцов и самок практически одинакова, что указывает на различные элементы контроля отдельных этапов генетической рекомбинации в половых и соматических клетках. В гетерохроматических районах, в частности прицентромерных, частота кроссинговера снижена, и поэтому истинное расстояние между генами в этих участках может быть изменено.
Обнаружены гены, выполняющие роль запирателей кроссинговера, но есть также гены, повышающие его частоту. Они иногда могут индуцировать заметное число кроссоверов у самцов дрозофилы. В качестве запирателей кроссинговера могут выступать также хромосомные перестройки, в частности инверсии. Они нарушают нормальную конъюгацию хромосом в зиготене.
Обнаружено, что на частоту кроссинговера влияют возраст организма, а также экзогенные факторы: температура, радиация, концентрация солей, химические мутагены, лекарства, гормоны. При большинстве указанных воздействий частота кроссинговера повышается.
В целом кроссинговер представляет собой один из регулярных генетических процессов, контролируемых многими генами как непосредственно, так и через физиологическое состояние мейотических или митотических клеток. Частота различных типов рекомбинаций (мейотический, митотический кроссинговер и сестринские, хроматидные обмены) может служить мерой действия мутагенов, канцерогенов, антибиотиков и др.
Законы наследования Моргана и вытекающие из них принципы наследственности. Огромную роль в создании и развитии генетики сыграли работы Т.Моргана. Он автор хромосомной теории наследственности. Им были открыты законы наследования: наследование признаков, сцепленных с полом, сцепленное наследование.
Из этих законов вытекает следующие принципы наследственности:
1. Фактор-ген есть определённый локус хромосомы.
2. Аллели гена расположены в идентичных локусах гомологичных хромосом.
3. Гены расположены в хромосоме линейно.
4. Кроссинговер – регулярный процесс обмена генами между гомологичными хромосомами.
Мобильные элементы генома. В 1948 г. американская исследовательница Мак-Клинток открыла у кукурузы гены перемещающиеся из одного участка хромосомы в другой и назвала феномен транспозицией, а сами гены контролириующими элементами (КЭ). 1.Эти элементы могут перемещаться из одного сайта в другой; 2. их встраивание в данный район влияет на активность генов расположенных рядом; 3. утрата КЭ в данном локусе превращает прежде мутабильный локус в стабильный; 4. в сайтах, в которых присутствуют КЭ, могут возникать делеции, транслокации, транспозиции, инверсии, а также разрывы хромосом. В 1983 г. за открытие мобильных генетических элементов Нобелевская премия была присуждена Барбаре Мак-Клинток.
Наличие мобильных элементов в геномах имеет разнообразные последствия:
1. Перемещения и внедрение мобильных элементов в гены может вызывать мутации;
2. Изменение состояния активности генов;
3. Формирование хромосомных перестроек;
4. Формирование теломер.
5. Участие в горизонтальном переносе генов;
6. Транспозоны на основе Р-элемента используют для трансформации у эукариот, клонирования генов, поиска энхансеров и т.д.
У прокариот существуют три типа мобильных элементов – IS-элементы (инсерции), транспозоны, и некоторые бактериофаги. IS-элементы встраиваются в любой участок ДНК, часто вызывают мутации, разрушая кодирующие или регуляторные последовательности, влияют на экспрессию соседних генов. Бактериофаг может вызывать мутации в результате встраивания.
Aллель - вариант (состояние) гена, локализованного в определенном локусе (месте) хромосомы.
Aллельные гены - гены, расположенные в одинаковых (идентичных) локусах гомологичных хромосом.
Аллели множественные - гены, представленные в популяции более чем в двух разных вариантах (состояниях). Механизм возникновения независимые генные мутации во дном локусе хромосомы.
Множественные аллели определяют варианты_ одного признака. Например, система групп крови АВО У человека кодируется тремя генами: Ja, Jb,i
Аутосомы - неполовые хромосомы, имеющие одинаковые размеры и форму у особей разных полов. У человека обозначаются цифрами от 1 до 22. Совокупности генов одной и той же аутосомы у разных индивидуумов отличаются комбинациями доминантных и рецессивных генов.
Гамета - половая клетка организма (яйцеклетка или сперматозоид).
Гаплоидный набор хромосом - определяется, как правило, в гаметах и содержит одну из каждой пары аутосом и одну половую хромосому (Х или У).
Гемизиготный генотип - генотип, в котором представлен только один аллельный ген. В норме, это характерно для генов, локализованных в негомологичных участках половых хромосом. В гемизиготном состоянии единственная аллель всегда проявляет себя в фенотипе.
Геном - совокупность генов всех особей определенного вида.
Генотип - совокупность всех генов диплоидной (соматической) клетки (в т.ч. митохондрий и пластид).
Генофонд - совокупность всех генов, определяющих у особей определенной популяции.
Гетерозиготный организм - особь, У которой в идентичных локусах гомологичных хромосом располагаются разные аллельные гены. При скрещивании гетерозиготных организмов происходит расщепление по генотипу и фенотипу в соответствии с законами Г.Менделя.
Гомозиготный организм - особь, у которой в идентичных локусax гомологичных хромосом располагаются одинаковые аллельные гены: оба доминантных (гомозиготный доминантный генотип) или оба рецессивных (гомозиготный рецессивный генотип).
Диплоидный набор хромосом - полный парный набор хромосом, содержащийся в соматических клетках (все клетки организма, за исключением половых).
Доминантный ген - ген, признак которого обычно проявляется у гетерозиготных организмов. Степень проявления доминантности зависит от формы взаимодействия аллельныx генов.
Доминирование полное - форма взаимодействия аллельных генов, при которой доминантный ген полностью подавляет действие рецессивного гена и фенотип гомозиготных доминантных и гетерозиготных организмов сходный.
Доминирование неполное - форма взаимодействия aллельных генов, при которой имеет место промежуточное проявление признака у гетерозиготных организмов по сравнению с гомозиготными. При этом степень проявления признака имеет следующую последовательность: АА > Аа > аа.
Кариотип - диплоидный набор хромосом, характеризующийся совокупностью признаков (число, форма, особенность дифференциального окрашивания). Кариотип является важнейшей цитогенетической характеристикой вида.
Кодоминирование - форма взаимодействия аллельных генов, при которой два разных доминантных аллельных генов проявляют себя. в фенотипе в равной степени. Пример, IV группу крови у человека определяет генотип JA J в.
Менделирующие признаки - наследственные признаки, которые контролируются аллеJ1!>НЫМИ генами и наследование их происходит в соответствии с законами моногибридного скрещивания Г.Менделя.
Наследование - способ передачи наследственной информации между поколениями. Варианты наследования зависят от локализации ДНК в структурных компонентах клетки. Различают аутосомное, X-сцепленное, голандрическое (У -сцепленное) и цитоплазматическое наследование.
Наследственность - общее свойство живых организмов обеспечивать структурную и функциональную преемственность между поколениями, а также специфический характер индивидуального развития.
Признак - любое свойство или качество (морфологическое биохимическое, иммунологическое, клиническое), которое отличает один организм от другого.
Фенотип - совокупность всех признаков организма.
Хромосомы половые - хромосомы, определяющие генетический пол организма - Х и У. У человека женский пол является гомогаметным – в яйцеклетках содержатся по одной Х-хромосоме (кариотип женщин 46,ХХ), а мужской пол является гетерогаметным - в сперматозоидах находится либо Х-хромосома, либо У-хромосома (кариотип мужчин 46,ХУ).
Хромосомы гомологичные - хромосомы, имеющие одинаковую ДЛИНУ: форму и характерные особенности дифференциального окрашивания. В диплоидном наборе содержатся 2 гомологичные хромосомы - аутосомы с 1 по 22 пары, у женщин - две Х-хромосомы. У мужчин половые хромосомы (Х и у) являются негомологичными.
1. Биология /Под редакцией В.Н.Ярыгина. в 2-х кн. М., Высшая школа, 2006. -кн. l, с. 61-65, 88-90, 115-125, 137-141, 155-158,222-227.
2. Лекционный материал
Темы учебно-исследовательской работы студентов: 1. Зарождение и становление генетики как науки. Научные труды
Г.Менделя, А.Вейсмана, х.де Фриза, В.Иоганнсена, Т.Моргана.
2. Генетические исследования в СССР.
3. Менделирующие признаки человека: норма и патология.
Прежде чем рассматривать вопросы взаимодействия генов необходимо ознакомиться с основными терминами и определениями, используемыми при изучении данного вопроса. Нам уже известно, что наследственные признаки обусловлены генами.
Гены – отдельные участки ДНК хромосом, ответственные за синтез одного белка.
Локус – место расположения гена на хромосоме.
В каждой паре гомологичных хромосом содержатся два родственных гена, которые отвечают за развитие одного признака
Локусы родственных генов расположены в одинаковых местах гомологичныххромосом
Аллель – один ген из пары, находящийся в сходном локусе гомологичных хромосом и контролирующий развитие альтернативных признаков. Аллелем называется также форма состояния гена.
Гомозиготные или чистые организмы – имеющие в одном и том же локусе гомологичных хромосом одинаковые по характеру действия гены (АА, аа, ВВ, вв).
Гетерозиготные или гибридные организмы – имеющие в одном и том же локусе гомологичных хромосом разные по характеру действия гены (Аа, Вв).
Решетка Пеннета – схематическое отображение процесса скрещивания.
Плейотропия – множественное действие гена, когда один ген ответственен за ряд фенотипических эффектов.
Полигенная детерминация – совместное действие нескольких генов на один признак.
Установленные Г. Менделем закономерности независимого расщепления признаковв потомстве гибридов распространяются на все случаи, когда каждый отдельный ген определяет развитие одного наследственного признака. Наряду с этим накоплены многочисленные факты, указывающие на сложные взаимодействия генов. Выяснилось, что один и тот же ген может влиять на несколько различных признаков и, наоборот, один и тот же наследственный признак развивается под влиянием многих генов. Известны два типа взаимодействия генов: аллельное и неаллельное.
Известны три основные формы взаимодействия между аллеломорфными генами: полное доминирование; неполное доминирование; и независимое проявление.
Полное доминирование наблюдается когда закономерности наследования подчиняются законам Менделя, когда в фенотипе гетерозигот присутствует продукт одного гена. При перекрестном опылении двух гомозиготных особей с генотипом АА и аа в первом гибридном поколении все растения по фенотипу будут одинаковы, а по генотипу будут гетерозиготными Аа.
Неполное доминирование – при котором фенотип гетерозигот имеет среднее значение между доминантными и рецессивными гомозиготами.
Простейший пример аллельного взаимодействия генов - неполное доминирование при скрещивании белых и красных цветков у львиного зева и получение в результате розовых цветков. Во втором гибридном поколении идет расщепление: одно красноцветковое растение, два с розовыми цветками и одно с белыми цветками. При этом наблюдается полное соответствие между фенотипом и генотипом - гомозиготы АА имеют красные цветки, гетерозиготы Аа - розовые и гомозиготы аа - белые. (составить решетку Пеннета).
Кодоминирование – взаимодействие аллельных генов, при котором у гетерозигот в фенотипе присутствует продукт обоих генов.
При кодоминировании у гетерозиготных организмов каждый из аллеломорфных генов вызывает формирование контролируемого им признака независимо от того, какой из других аллеломорфных ему сопутствует. Примером кодоминирования является наследования у человека группы крови систем АВО. Группа крови контролируется серией множественных аллелей одного гена. Три аллели формируют шесть генотипов ОО – первая, АА или АО – вторая, ВВ или ВО – третья, АВ – четвертая группа крови.
Взаимодействие неаллеломорфных генов:
Гены расположенные в разных локусах и ответственные за проявление одного гена называются неаллельными.
Известны четыре формы взаимодействия: 1) комплектарность , при которой соответствующий признак развивается только в присутствии двух определенных неаллеломорфных генов; 2) эпистаз , при котором один из генов полностью подавляет действие другого неаллеломорфного гена; 3) полимерия, при которой неаллеломорфные гены действуют на формирование одного и того же признака и вызывают примерно одинаковые его изменения; 4) модификация, при которой одни гены видоизменяют действие других, подавляя, интенсифицируя или ослабляя их.
Комплементарные гены – обуславливающие при совместном сочетании новое фенотипическое проявление признака. Расщепление – 9:3:3:1, 9:7, 9:3:4, 9:6:1
9:3:3:1 – каждый доминантный ген имеет самостоятельное фенотипическое проявление, сочетание в генотипе этих двух генов обуславливает новое фенотипическое проявление, а их отсутствие – не дает развитие признака. Например – ген А обуславливает развитие голубой окраски оперения волнистых попугайчиков, ген В – желтой, а попугайчики с генотипом А_В_ – имеют зеленую окраску, а с генотипом ааbb – белую
9:7 – доминантные и рецессивные аллели комплементарных генов не имеют самостоятельного фенотипического проявления. Например, пурпурная окраска венчика цветка у душистого горошка развивается только при совместном сочетании в генотипе доминантных генов А и В, во всех остальных случаях окраска отсутствует, и венчик оказывается белым
9:3:4 – доминантные и рецессивные аллели комплементарных генов имеют самостоятельное фенотипическое проявление. Например окраска у кроликов определяется двумя комплементарными генами: А – наличие окраски, а – отсутствие, В – черная окраска, b – голубая окраска
9:6:1 – сочетание доминантных аллелей комплементарных генов обеспечивает формирование одного признака, сочетание рецессивных аллелей этих генов – другого, а наличие в генотипе только одного из доминантных генов – третьего. Например, тыквы с генотипом А_В_ имеют дисковидную форму плода, с генотипом ааbb – удлиненную, а с генотипом А_bb или ааВ_ - сферическую
Эпистаз – взаимодействие неаллельных генов, при котором один из них подавляет действие другого. Ген, подавляющий действие другого неаллельного гена, называется супрессором или ингибитором, и обозначается I или S. Подавляемый ген называется гипостатичным. Эпистаз может быть доминантным и рецессивным.
Доминантным эпистазом называют подавление действия гена доминантной аллелью другого гена.
Расщепление: 13:3 – наблюдается в том случае, если доминантная аллель эпистатического гена не имеет своего фенотипического проявления, а лишь подавляет действие другого гена, в то время как его рецессивная аллель не влияет на проявление признака. Например, у некоторых пород кур наличие доминантного эпистатического гена подавляет развитие окраски оперения, при его отсутствии куры окрашены
12:3:1 – наблюдается в том случае, если гомозиготная по рецессивным признакам особь имеет особый фенотип. Например, от скрещивания двух гетерозиготных собак щенки с генотипом I_аа имеют белую окраску, а с генотипом iiА_ – черную, а с генотипом iiаа – коричневую
Взаимодействие неаллельных генов, при котором рецессивная аллель эпистатического гена в гомозиготном состоянии подавляет действие другого гена называется рецессивным эпистазом. При одинарном рецессивном эпистазе рецессивная аллель одного гена подавляет действие другого (аа подавляет В_). При двойном – рецессивная аллель каждого гена в гомозиготном состоянии подавляет действие доминантной аллели (аа подавляет В_, bb подавляет А_).Расщепление 9:3:4 или 9:7
Полимерия – взаимодействие неаллельных генов, однозначно влияющих на развитие одного и того же признака.
Такие гены называются полимерными или множественными и обозначаются одинаковыми буквами с соответствующими индексом (А1,А2,А3). Чаще всего полимерные гены контролируют количественные признаки (высота, масса, и т.д.)
Полимерия может быть кумулятивной (суммирующей, аддитивной) и некумулятивной
При кумулятивной полимерии степень проявления признака зависит от числа доминантных аллелей соответствующих полимерных генов. Например, чем больше доминантных аллелей генов, отвечающих за окраску кожи, содержится в генотипе человека, тем его кожа темнее
При некумулятивной полимерии степень развития признака зависит не от количества доминантных аллелей, а лишь от их наличия в генотипе. Например, куры с генотипом а1,а2,а3 имеют неоперенные ноги, во всех остальных случаях – ноги оперены
Модифицирующие гены - гены, усиливающие или ослабляющие действие других генов. Сами гены-модификаторы собственного проявления не имеют. Теоретически любой ген, взаимодействуя с остальными, должен модифицировать проявление другого гена. Однако существуют группы генов, которые отчетливо показывают свое модифицирующее действие на проявление несколько генов. У таких генов-модификаторов часто не обнаруживается их самостоятельное действие на особь. Об их существовании мы узнаем по их влиянию на другие гены. По типу своего действия гены модификаторы представлены двумя категориями: 1) гены, усиливающие проявление признака, детерминируемого другим геном; 2) гены, ослабляющие действие другого гена.
Сцепленное наследование.
Третий закон Менделя - правило независимого наследования имеет существенное ограничение. Он действителен только в тех случаях, когда гены локализованы в разных хромосомах. Когда же неаллеломорфные гены располагаются в одной хромосоме в линейном порядке, независимого расщепления не наблюдается, а наблюдается совместное наследование генов, ограничивающее их свободное комбинирование, Т. Морган назвал это явление сцеплением генов или сцепленным наследованием. Если по теории Менделя при скрещивании АВ и ав получается гибрид АаВв, образующий четыре сорта гамет АВ, Ав, Ва, ва. В соответствии с этим в анализирующем скрещивании осуществляется расщепление 1:1:1:1 т.е. по 25 %. Однако по мере накопления фактов наблюдается отклонения от такого расщепления. В отдельных случаях новые комбинации Ав и Ва совсем отсутствовали - наблюдалось полное сцепление между генами исходных форм, которое проявлялось в равных количествах - по 50 %. Гены чаще наследовались в исходном состоянии (были сцеплены).
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, одна хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков.
Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался выдающийся американский генетик Т.Морган.
Явление совместного наследования признаков называют сцепленным . Материальной основой сцепления генов является хромосома. Гены, локализованные в одной хромосоме, наследуются совместно и образуют одну группу сцепления. Количество групп сцепления равно гаплпоидному набору хромосом. Явление совместного наследования генов, локализованных в одной хромосоме, называют сцепленным наследованием. Сцепленное наследование генов, локализованных в одной хромосоме, называют законом Моргана.
Различают два варианта локализации доминантных и рецессивных аллелей генов, относящихся к одной группе сцепления:
Цис-положение , при котором доминантные аллели находятся в одной из пары гомологичных хромосом, а рецессивные – в другой.
Транс-положение , при котором доминантные и рецессивные аллели гена находятся в разных гомологичных хромосомах.
Гены в хромосомах имеют разную силу сцепления. Сцепление может быть полным – если гены, относящиеся к одной группе сцепления, всегда наследуются вместе; неполным, если между генами, относящимися к одной группе сцепления, возможна рекомбинация.
Исследования Т. Моргана показали, что в гомологичной паре хромосом частично происходит обмен генами. Процесс обмена был назван кроссинговером. Кроссинговер способствует новому сочетанию генов, находящихся в гомологичных хромосомах и тем самым увеличивает роль комбинативной изменчивости в эволюции. Изучение явления кроссинговера, нарушающего сцепление генов, утвердило представление о строго фиксированном расположении генов вдоль хромосом. Принцип линейного расположения известен как второй закон Т.Моргана.
Сцепление генов может нарушаться в процессе кроссинговера ; это приводит к образованию рекомбинантных хромосом. В зависимости от особенностей образования гамет, различают:
кроссоверные гаметы – гаметы с хромосомами, претерпевшими кроссинговер;
некроссоверные гаметы – гаметы с хромосомами, образованными без кроссинговера.
При сцепленном наследовании признаков, гены которых локализованы в одной хромосоме, соотношение фенотирических классов потомства, полученного от скрещивания, часто отличается от классического менделеевского. Это связано с тем, что часть гамет родительских особей является кроссоверной, а часть – некроссоверной.
Вероятность возникновения перекреста между генами зависит от их расположения в хромосоме: чем дальше друг от друга расположены гены, тем выше вероятность перекреста между ними. За единицу расстояния между генами, находящимися в одной хромосоме, принят 1 % кроссинговера. Его величина зависит от силы сцепления между генами и соответствует проценту рекомбинантных особей (особей, образованных с участием кроссоверных гамет) от общего числа потомков, полученных при скрещивании. В честь Т.Моргана единица расстояния между генами названа морганидой.
Процент кроссинговера между генами вычисляют по формуле:
Х = (а+в) х 100
П
где Х – процент кроссинговера, а – число кроссоверных особей одного класса, в – число кроссоверных особей другого класса, п – общее число особей, полученных от анализирующего скрещивания.
Величина кроссинговера не превышает 50 % , если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Согласно хромосомной теории наследственности, гены в хромосомах располагаются линейно. Генетическая карта хромосомы – схематическое изображение относительного положения генов, входящих в одну группу сцепления.
О положении гена в группе сцепления судят по проценту кроссинговера (количеству кроссоверных особей): чем больше процент кроссинговера или количество кроссоверных особей в F a , тем дальше будут расположены анализируемые гены/
Задачи на сцепленное наследование решаются аналогично задачам на моно- и дигибридное скрещивание. Однако при сцепленном наследовании гены, контролирующие развитие анализируемых признаков, локализованы в одной хромосоме. Поэтому наследование этих признаков не подчиняется законам Менделя.
Генотипы скрещиваемых особей и гибридов следует писать в хромосомной форме;
При записи генотипов следует учитывать расположение генов в хромосомах гомологичной пары (цис- или транс-положение). При цис-положении доминантные аллели генов находятся в одной хромосоме, а рецессивные – в другой. При транс-положении в хромосоме располагаются доминантная аллель одного гена и рецессивная – другого.
При полном сцеплении особь, гетерозиготная по всем рассматриваемым признакам, образует два типа гамет
При неполном сцеплении происходит образование кросоверных и некроссоверных гамет.
Количество некроссоверных гамет всегда больше, чем кроссоверных;
Организм всегда образует равное количество разных типов как кроссоверных, так и некроссоверных гамет;
Процентное соотношение кроссоверных и некроссоверных гамет зависит от расстояния между генами;
Если известно расстояние между генами (в процентах кроссинговера или морганидах), то количество кроссоверных гамет определенного типа можно вычислить по формуле
П = % кроссинговера (2)
где -количество кроссоверных гамет определенного типа;
Если известно количество кроссоверных особей, то процент кроссинговера между генами вычисляют по формуле (1)
Если рассматриваются признаки, гены которых входят в состав разных групп сцепления, то вероятность объединения генов разных групп сцепления в одной гамете равна произведению вероятностей каждого гена, образующего эту гамету.
Чтобы определить вероятность появления разных сортов зигот, надо перемножить частоты гамет, образующих эту зиготу.
У левкоев махровость поддерживается одним рецессивным геном s , доминирует простой цветок S . Доминантный аллель S сцеплен с рецессивным аллелем l , который вызывает отмирание пыльцы, а рецессивный s с доминантным L - нормально развитой пыльцой
У петуний неполное доминирование махровости, так что гетерозиготы Gg образуют слабо махровые цветки по сравнению с гомозиготами GG , дающими растение с махровыми цветками.
Имеется еще ген А - усилитель махровости, который действует не самостоятельно при наличии его (АА или Аа ) удается различать густо и слабомахровые растения, при отсутствии (аа )наблюдается моногибридное расщепление 3:1
У гвоздики турецкой махровость определяется одним рецессивным геном, вызывающим одновременно мужскую стерильность. Так как махровые растения могут служить лишь материнской формой, 100%-ая махровость в потомстве неосуществима.
Теоретически возможно получение 50%-ной при опылении махровых растений (ее ) простыми (Ее
У гвоздики садовой (формы Шабо, Маргарита, Венская, Гренадин) махровость определяется одним геном с неполным доминированием. Гомозиготы GG отличаются большей степенью махровости и почти полной мужской стерильностью. Гетерозиготы Gg образуют менее махровые цветки.
Наследование махровости редко бывает одинаковым и постоянным у различных сортов и форм одного и того же вида и тем более семейства. Особенно неустойчива она у видов и сортов семейства сложноцветных и в большей степени зависит от уровня агротехники и погодных условий.
Махровость у сложноцветных определяется несколькими или многими генами, большая часть которых имеет доминантное действие
Это относится к астрам, ноготкам, бархатцам и маргариткам.
Для поддержания махровости необходим постоянный массовый отбор махровых растений при сохранении достаточного количества растений с диском обоеполых трубчатых цветков в центре соцветия как источника пыльцы.